Le cerveau est un processeur fractal massivement parallèle qui génère des modèles de champs électromagnétiques spatiotemporels complexes en corrélation avec la cognition et la perception. Une propriété clé d’un système fractal est la complexité sans échelle, ce qui signifie que le degré de complexité du système est invariant sous l’effet de l’échelle – par exemple, en utilisant une quantification en loi de puissance, on peut montrer que le degré de complexité du cerveau humain est approximativement invariant du niveau tissulaire au niveau cellulaire et au niveau moléculaire. Les potentiels de champ électrique et les réponses à la résonance magnétique du cerveau présentent tous une dynamique sans échelle [1], et cette dynamique cérébrale invariante contient des structures spatio-temporelles complexes qui sont modulées par l’exécution d’une tâche. Des études antérieures ont montré que des corrélations temporelles sans échelle relient les modes vibratoires des neurones individuels aux biopolymères de type antennes fractales (résonateurs diélectriques), tels que les microtubules, à l’intérieur de tout neurone individuel, démontrant une liaison sans échelle des résonances diélectriques du niveau cellulaire au niveau moléculaire dans les neurones cérébraux [2].
Des expériences récentes menées par le groupe de recherche de Bandyopadhyay ont permis de mesurer directement une paire de vortex de champ électrique générés par le réseau quadrillé de filaments intermédiaires d’actine situé juste sous la membrane neuronale, qui active de manière cohérente tous les canaux ioniques dans une zone circulaire de la bicouche lipidique de la membrane lors de la propagation d’un signal électrique [3]. Les images de résonance diélectrique obtenues par le groupe de recherche ont montré que le réseau ordonné de microtubules au cœur du corps cellulaire du neurone initie et commande un réseau cylindrique superstructurel en forme de grille de filaments intermédiaires de bêta-spectrine et d’actine qui s’interfacent avec la bicouche lipidique du neurone pour générer une paire de vortex électriques, qui à leur tour régulent la synchronisation du signal de vortex électrique-ionique de la membrane neuronale. Ces résultats s’inscrivent dans la continuité de la caractérisation empirique et de la démonstration par le groupe de recherche de Bandyopadhyay du rôle direct des microtubules et du réseau filamentaire subcellulaire associé dans la dynamique électromagnétique sans échelle qui sous-tend la cognition et la perception au sein du cerveau [4], fonctionnant comme des guides d’ondes circulaires et des résonateurs [5], ainsi que comme des processeurs d’information [6].
En utilisant la technologie des sondes coaxiales, avec une résolution à l’échelle nanoscopique, et la microscopie à résonance diélectrique, le groupe de recherche de Bandyopadhyay a pu étudier les communications ultrarapides de signaux électromagnétiques provenant du faisceau filamentaire de microtubules à l’intérieur de neurones uniques vers un réseau de type grille d’actine situé juste en dessous de la membrane neuronale qui régule la synchronisation des potentiels d’action. Les chercheurs ont constaté que les transmissions filamentaires saturaient 200 microsecondes avant le déclenchement d’un potentiel d’action par le neurone, un domaine temporel 1000 fois plus rapide que la pointe nerveuse. Les mesures effectuées à l’aide de sondes coaxiales à l’échelle nanométrique dans trois domaines temporels simultanés (millisecondes, microsecondes et nanosecondes) ont révélé que les transmissions électromagnétiques à travers les protéines tubulines, les microtubules et les neurones présentaient un profil temporel similaire, sous la forme d’un triplet de triplets sans échelle (film 1, figure 2).
Film 1. Roue de fréquence pour le motif de fréquence de résonance triplet-triplet pour la protéine de tubuline de 4-4 nm de large, pour le nanofil de microtubule de 25 nm de large et le segment initial d’axone de 1-μm de large d’un neurone. Reproduit de K. Saxena et al, « Fractal, Scale Free Electromagnetic Resonance of a Single Brain Extracted Microtubule Nanowire, a Single Tubulin Protein and a Single Neuron, » Fractal and Fractional, vol. 4, no. 2, Art. no. 2, Jun. 2020, [2].
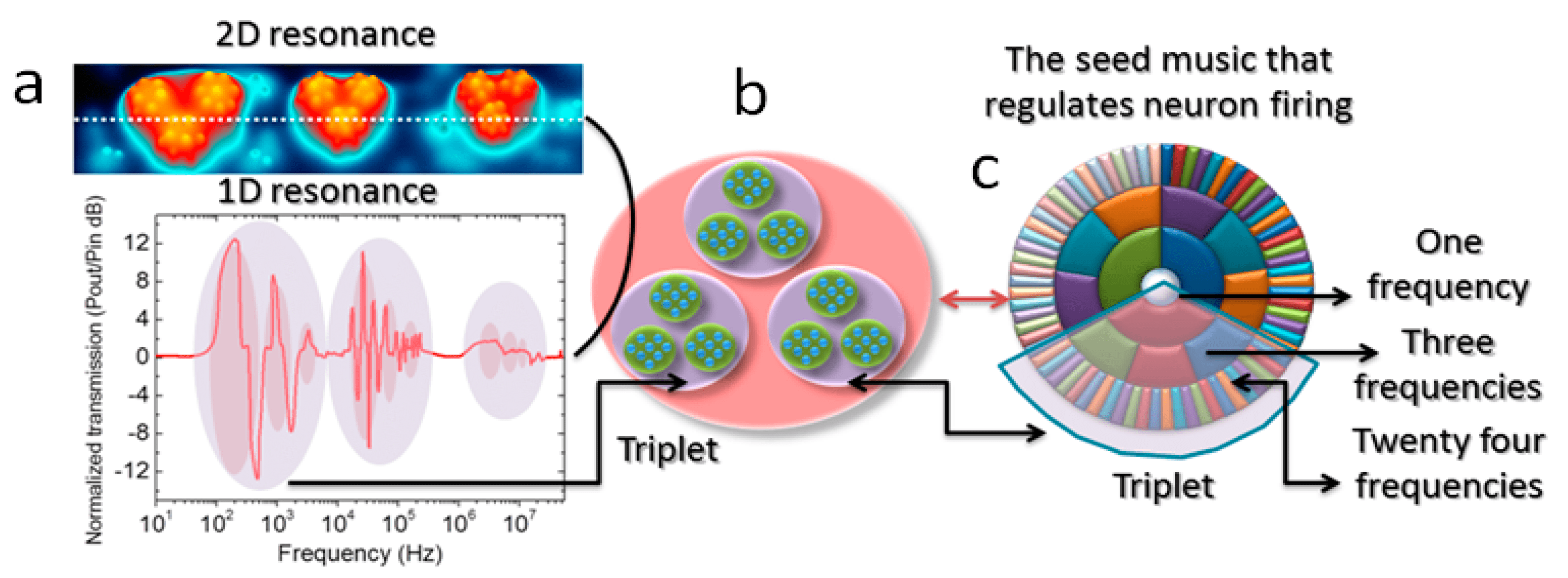
Figure 1. (a) Résonance 2D d’un microtubule isolé, montrant une bande triplet de 10-300 kHz, 10-230 MHz et 1-20 GHz. Une ligne pointillée indique l’emplacement de la mesure de la résonance 1D (en haut), les données sont indiquées sous le panneau (a). Le tracé de résonance 3D du panneau a est représenté par un nid de neuf cercles (neuf cercles à l’intérieur de trois cercles à l’intérieur d’un cercle = 13 cercles). Un triplet dans un diagramme de résonance 1D du panneau est relié par une ombre et une flèche (a), au schéma du panneau (b) ; (c) Le diagramme circulaire triplet-triplet du panneau b est une réplique des données expérimentales de résonance (panneau a), cependant, les oscillations de résonance suivent une condition périodique. Si des limites périodiques sont appliquées, le panneau b ressemble au panneau c. On peut trouver le triplet dans le panneau c, chaque triplet a une seule fréquence, et sa période totale est la somme des trois fréquences à l’intérieur : chacune des trois a des périodes de huit fréquences (un cas particulier de tubuline). Reproduit de K. Saxena et al, « Fractal, Scale Free Electromagnetic Resonance of a Single Brain Extracted Microtubule Nanowire, a Single Tubulin Protein and a Single Neuron, » Fractal and Fractional, vol. 4, no. 2, Art. no. 2, Jun. 2020, [2].
Film 2. Triplet de bandes de résonance triplet dans un microtubule, animation montrant les modes vibratoires complexes.
En utilisant l’imagerie par résonance diélectrique, les chercheurs ont observé des sections du noyau neurofilament-microtubule, qui ont acquis une énergie électromagnétique de résonance et, en raison de la résonance diélectrique et des symétries locales des filaments orientés en hélice, ont sélectivement transporté l’énergie quantifiée à travers le réseau subcellulaire, contribuant ainsi au potentiel de champ électrique endogène du neurone. Ensuite, en faisant résonner sélectivement la superstructure de la grille cylindrique bêta-spectrine-actine avec la fréquence de résonance appropriée, les chercheurs ont observé la formation d’anneaux brillants de tourbillons électromagnétiques juste sous la membrane plasmique (figure 2). Fait important, l’équipe de recherche décrit la topologie précise du potentiel d’action qui se propage comme un anneau de champs électriques autour du périmètre circulaire de la branche axonale, se propageant donc comme un paquet d’ondes gaussiennes en 3D. Cela contraste avec le modèle neuronal conventionnel qui considère uniquement les transmissions neuronales comme des potentiels de pointe 1D, ce qui, comme le souligne le groupe de recherche, ne correspond pas à ce qui se passe dans le réseau dans un scénario réel. Comme l’observe directement la méthodologie avancée de l’équipe de recherche à une résolution nanométrique, l’influx nerveux est un vortex électrique-ionique qui couvre le périmètre des branches axonales ou dendritiques et qui est régulé par des signaux électromagnétiques provenant du réseau filamentaire subcellulaire sous-jacent.
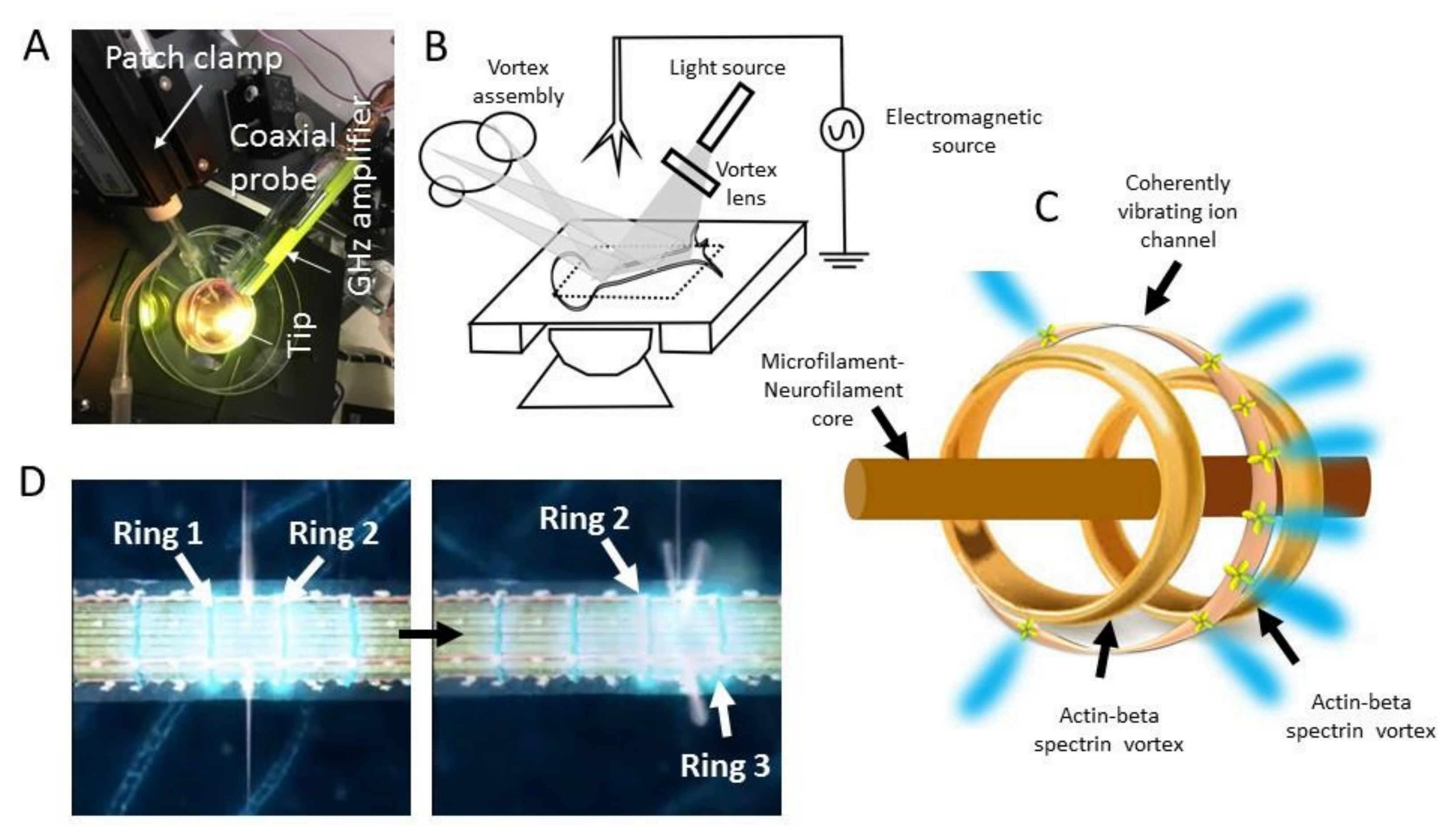
Figure 2 : (A) Montage expérimental pour des mesures combinées de patch-clamp et de sonde coaxiale sur une plaque de culture. La sonde atomique coaxiale récemment inventée détecte plusieurs domaines temporels à la fois, même en dessous de 0,5 nm. Sa cavité est située entre une aiguille en métal Pt et un cylindre en métal Au et piège les vibrations (10-20 watts) dans son voisinage, la géométrie conique de la cavité amplifiant ce signal. Le résonateur diélectrique intégré (verre) augmente le facteur Q (~105) à toutes les fréquences que sa géométrie conique permet (3 kHz à 40 GHz). Contrairement au patch-clamp, une sonde atomique lit quatre signaux simultanément, filtre le bruit, mesure quatre vibrations distinctes de complexes protéiques, sans bruit (S/N~105), à l’intérieur d’un neurone. (B) Montage expérimental pour la combinaison simultanée de micro-ondes et d’ondes radio avec l’optique quantique. Un laser monochromatique (633,5 nm) passe à travers une lentille vortex pour générer un vortex optique, qui est projeté sur la région de l’axone. Une caméra à semi-conducteur prend des images de l’ensemble des tourbillons réfléchis ou transmis. (C) Schéma du cœur du neurofilament, transportant l’énergie EM résonnante vers la superstructure filamentaire intermédiaire actine-bêta-spectrine, qui active de manière cohérente les canaux ioniques membranaires pour contrôler le temps de déclenchement ionique précis. (D) Deux panneaux montrent qu’au cours de la première étape, les anneaux 1 et 2 de la bêta-spectrine-actine sont actifs, les ions étant libérés dans la voie circulaire qui les relie. Ensuite, l’anneau 1 s’éteint, les anneaux 2 et 3 sont actifs. Les canaux ioniques situés entre eux sont activés pour la libération d’ions. Reproduit de P. Singh et al, « Cytoskeletal Filaments Deep Inside a Neuron Are not Silent : They Regulate the Precise Timing of Nerve Spikes Using a Pair of Vortices », Symmetry, vol. 13, no. 5, Art. no. 5, mai 2021, [3].
Le groupe de recherche Bandyopadhyay a décrit l’importance de la synchronisation, ou de la fréquence, des signaux électriques neuro-filamentaires dans la génération des modèles de champs électromagnétiques spatiotemporels spécifiques et hautement structurés du cerveau. En fait, cela a conduit les chercheurs à proposer un modèle de cristal de temps auto-opérant du cerveau humain, et ils ont discuté de la possibilité de reproduire complètement la fonctionnalité cognitive du cerveau par le biais d’une architecture fractale 3D composée uniquement d’horloges [7]. Cela souligne l’importance des dernières découvertes de l’équipe, car les tourbillons électromagnétiques générés dans le réseau sous-membranaire d’actine produisent une horloge à la microseconde qui régule la conduction ionique à la milliseconde de l’axone neuronal. Cette découverte corrobore des recherches antérieures dont nous avons rendu compte, telles que le traitement à plusieurs niveaux sous-seuil dans les dendrites, dans lequel l’architecture subsynaptique complexe confère à un seul neurone la puissance de calcul normalement attribuée à des réseaux neuronaux multicouches entiers [8].
L’équipe de recherche de Bandyopadhyay décrit les oscillations périodiques fractalisées des microtubules à l’actine et aux membranes neuronales comme des horloges interconnectées, l’ordre et l’arrangement des horloges révélant une symétrie dans la structure d’information du neurone. L’assemblage de vortex électromagnétiques/optiques est une structure géométrique et en les reliant à différentes symétries de l’architecture neuronale, une perspective matérielle a été ajoutée aux observations du processus de signalisation collective dans les circuits filamentaires. Les chercheurs affirment qu’à l’avenir, ils mèneront des recherches pour mieux comprendre comment les mécanismes d’interaction et d’ajustement temporel de l’architecture de l’horloge fractale expliquent l’entropie et les réponses thermodynamiques d’un neurone. Grâce aux progrès de la technologie de la sonde coaxiale et de l’imagerie par résonance diélectrique, ils espèrent obtenir une meilleure résolution des sous-composants du neurone afin de faire correspondre chaque ensemble de fréquences de résonance à un sous-composant spécifique, ce qui permettra de mieux délimiter la dynamique de sous-seuil qui régule le potentiel électrique de la cellule.
La science unifiée en perspective
La complexité sans échelle associée au système biologique en général, et au neurone en particulier, signifie qu’à l’intérieur de chaque cellule se trouve un véritable cerveau macromoléculaire, au moins en termes de complexité structurelle et peut-être aussi, dans une certaine mesure, de complexité fonctionnelle – une architecture fractale interconnectée. Cela signifie que la vision extrêmement simpliste de la synapse en tant que bit numérique unique déforme la réalité de la situation. En effet, si nous devions utiliser le langage du modèle neuro-informatique, chaque « unité de calcul » contiendrait un véritable cerveau macromoléculaire. Il n’existe pas encore de technologie informatique ou humaine équivalente.
William Brown, Resonance Academy Big Questions Course, Lesson III : The Cellular Hologramic Information Nexus | Sentience and Memory Encoding in Cellular and Macromolecular Systems. 2018.
Les modèles complexes de champs électromagnétiques spatiotemporels, qui forment macroscopiquement des formes d’ondes continues, alors que microscopiquement ils sont composés d’oscillations électroniques discrètes et de tourbillons électromagnétiques, sont une indication directe de la façon dont le fonctionnement du cerveau et de la cognition elle-même est de nature semblable à celle d’un champ. Les comportements des champs, comme le champ électromagnétique, contiennent une sorte de qualité non locale inhérente, c’est pourquoi ils expliquent les forces physiques et l’action apparente à distance. Si l’on considère l’état de vide du champ électromagnétique, il est sans ambiguïté non local, avec une corrélation spatiale intrinsèque qui relie les systèmes indépendamment de la séparation spatiale ou temporelle (voir notre article Protocole de téléportation de l’énergie quantique). Cela signifie que notre esprit est semblable à un champ et qu’il fonctionne beaucoup plus comme la résonance morphique de Rupert Sheldrake que comme le modèle neuro-informatique conventionnel (Rupert Sheldrake a documenté expérimentalement des preuves de l’existence de signaux non locaux dans le cerveau et au cours de la cognition animale).
Le fonctionnement du cerveau à la manière d’un champ ne pourrait être plus évident que si l’on considère la biophysique des antennes fractales que l’on trouve à de nombreux niveaux du système neuronal, des sous-régions cérébrales entières aux neurones individuels, en passant par les composants neuronaux subcellulaires. Nombre de ces structures sont des résonateurs diélectriques et sont maintenant documentées empiriquement comme des guides d’ondes électromagnétiques. Ce sont ces systèmes qui couplent les fonctions biologiques au champ, qui est lui-même composé de résonateurs harmoniques dotés de fonctions d’hystérésis et de mémoire – raison pour laquelle nous appelons le tissu de l’espace l’espace-mémoire.
La mémoire spatiale est le siège des processus intelligents dans la nature et, en raison de sa vaste interconnectivité et de sa non-localité dans l’espace et le temps, c’est un domaine d’hyperintelligence naturelle. Il n’est donc pas surprenant que les structures de traitement de l’information du système biologique soient spécifiquement couplées au champ, puisque le système biologique a émergé et évolué avec le réseau hyperintelligent du champ. Même les tourbillons électromagnétiques décrits par l’équipe de recherche de Bandyopadhyay en témoignent, car la dynamique du tore est une caractéristique essentielle de la géométrie fondamentale de l’espace-temps, de sorte qu’un tourbillon électromagnétique à l’intérieur du neurone induit une dynamique toroïdale du plasma de Planck, pour une transmission d’énergie cohérente et une synchronisation collective maximales.
References
[1] B. J. He, “Scale-Free Properties of the Functional Magnetic Resonance Imaging Signal during Rest and Task,” J. Neurosci., vol. 31, no. 39, pp. 13786–13795, Sep. 2011, doi: 10.1523/JNEUROSCI.2111-11.2011.
[2] K. Saxena et al., “Fractal, Scale Free Electromagnetic Resonance of a Single Brain Extracted Microtubule Nanowire, a Single Tubulin Protein and a Single Neuron,” Fractal and Fractional, vol. 4, no. 2, Art. no. 2, Jun. 2020, doi: 10.3390/fractalfract4020011.
[3] P. Singh et al., “Cytoskeletal Filaments Deep Inside a Neuron Are not Silent: They Regulate the Precise Timing of Nerve Spikes Using a Pair of Vortices,” Symmetry, vol. 13, no. 5, Art. no. 5, May 2021, doi: 10.3390/sym13050821.
[4] S. Sahu, S. Ghosh, K. Hirata, D. Fujita, and A. Bandyopadhyay, “Multi-level memory-switching properties of a single brain microtubule,” Applied Physics Letters, vol. 102, no. 12, p. 123701, Mar. 2013, doi: 10.1063/1.4793995.
[5] F. Jelínek and J. Pokorný, “Microtubules in Biological Cells as Circular Waveguides and Resonators,” Electro- and Magnetobiology, vol. 20, no. 1, pp. 75–80, Jan. 2001, doi: 10.1081/JBC-100103161.
[6] T. J. A. Craddock, C. Beauchemin, and J. A. Tuszynski, “Information processing mechanisms in microtubules at physiological temperature: Model predictions for experimental tests,” Biosystems, vol. 97, no. 1, pp. 28–34, Jul. 2009, doi: 10.1016/j.biosystems.2009.04.001.
[7] P. Singh et al., “A Self-Operating Time Crystal Model of the Human Brain: Can We Replace Entire Brain Hardware with a 3D Fractal Architecture of Clocks Alone?,” Information, vol. 11, no. 5, Art. no. 5, May 2020, doi: 10.3390/info11050238.
[8] A. Gidon et al., “Dendritic action potentials and computation in human layer 2/3 cortical neurons,” Science, vol. 367, no. 6473, pp. 83–87, Jan. 2020, doi: 10.1126/science.aax6239.


