Synopsis Essentiel
Une analyse des effets photophysiques et photochimiques dans les méga-réseaux de tryptophane dans les architectures biologiques a révélé une cohérence collective entraînant une amélioration optique quantique, connue sous le nom de superradiance, et une interactivité à longue portée, même dans l’environnement thermique ambiant de la cellule [1]. De tels effets optiques quantiques collectifs dans les méga-réseaux de tryptophane peuvent sous-tendre des processus cellulaires clés d’orchestration et de signalisation, comme le transfert d’informations potentiellement ultrarapide dans les axones neuronaux, qui possèdent des faisceaux filamentaires de microtubules hautement ordonnés, enrichis de groupements de tryptophane coordonnés qui sont impliqués dans le traitement de l’information. Ces preuves empiriques appuient des théories telles que le réseau unifié de mémoire spatiale de Haramein et Brown [2] ou la réduction objective orchestrée de Hameroff et Penrose [3], qui impliquent des assemblages macromoléculaires subcellulaires tels que les mitochondries et les microtubules (la matrice réticulaire mitochondriale).
Ce qui a été trouvé
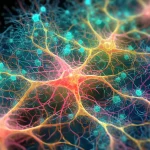
Une équipe de recherche internationale (des États-Unis, du Mexique, de la Suisse et de l’Italie) a réalisé une analyse complète combinant la modélisation informatique, l’analyse théorique et les mesures expérimentales pour documenter et caractériser le traitement collectif des signaux optiques dans les méga-réseaux de tryptophane tels que ceux que l’on trouve dans les microtubules. Par exemple, une super-architecture de microtubules, appelée centriole (figure 1), d’une longueur de 320 nm contient environ 105 chromophores de tryptophane (un chromophore est une molécule qui absorbe et émet de la lumière). À une telle taille, la résonance à longue portée et la synchronisation des oscillations dipolaires font de ces architectures protéiques des méga-réseaux et contribuent à expliquer des études antérieures comme celle de Bandopahdyay et al. [4] qui ont trouvé une coordination significative entre des microtubules uniques se comportant comme si l’ensemble de l’assemblage supramoléculaire était dans un état quantique, comme un condensat de Frohlich [5-8].
Cette découverte vient donc étayer les théories selon lesquelles les microtubules jouent un rôle essentiel dans le traitement de l’information cellulaire : ils pourraient jouer un rôle important dans les processus cognitifs tels que ceux qui impliquent les neurones. La réponse optique collective des réseaux de tryptophane dans les architectures protéiques est caractérisée par les spectres d’absorption et d’émission des chromophores du réseau. L’analyse de cette réponse optique collective des réseaux de tryptophane a permis d’identifier des améliorations superradiantes, un effet quantique qui se traduit par une intensité d’émission plus élevée d’un flux électromagnétique cohérent, qui pourrait être utilisé par la cellule pour la signalisation et le transport d’énergie à haut rendement.
La réponse optique collective peut être affectée par le désordre structurel, mais les grands réseaux de résidus polymères tryptophanes coordonnés examinés dans cette étude tendent à présenter une robustesse coopérative au désordre. Ce phénomène implique qu’une collection de chromophores tryptophanes identiques émettent spontanément un rayonnement cohérent intense. Les dipôles de transition des chromophores de tryptophane se synchronisent de manière cohérente, ce qui conduit à des états superradiants qui améliorent le transfert d’excitation au sein du réseau. Cet effet quantique coopératif se traduit par un pic d’intensité d’émission plus élevé que celui d’un seul chromophore. Ainsi, les propriétés photoexcitoniques collectives des grands réseaux de tryptophane dans les structures protéiques allant des dimères de tubuline individuels et des segments de microtubules aux super-architectures de microtubules telles que le centriole et les faisceaux neuronaux sont évidentes dans le rendement fluorescent mesuré, qui augmente de manière significative des tubulines individuelles aux assemblages de tubulines, une caractéristique clé de la superradiance et donc des propriétés cohérentes quantiques collectives.
La réponse optique collective est influencée par le couplage entre les chromophores tryptophanes induit par le champ électromagnétique. La réponse optique collective des réseaux de tryptophane dans les architectures de protéines est caractérisée par les améliorations du rendement quantique (QY). Le QY est un chiffre de mérite pour la coopérativité et une mesure robuste de l’amélioration de la réponse optique. Le QY n’est pas affecté de manière significative par le désordre statique, même pour des forces de désordre de la même magnitude que l’énergie à température ambiante. Cette robustesse est attribuée à la distribution de la force dipolaire superradiante parmi d’autres états excitoniques, tandis que les états proches de l’état superradiant en énergie présentent encore la majeure partie de la force dipolaire.
Par conséquent, la réponse optique collective des réseaux de tryptophane dans les architectures de protéines est caractérisée par une amélioration persistante du rendement quantique, même dans des conditions ambiantes. La réponse optique collective des réseaux de tryptophane dans les architectures protéiques est la formation d’états fortement superradiants dus à des interactions collectives entre des arrangements organisés de plus de 105 dipôles de transition de tryptophane excités par les UV dans les architectures de microtubules. Cela conduit à une amélioration du rendement quantique de la fluorescence, qui a été confirmée par des expériences. Le comportement superradiant du rendement quantique de la fluorescence est observé dans les structures de tubuline organisées de manière hiérarchique, telles que les microtubules, les centrioles et les faisceaux neuronaux. Le rendement quantique augmente dans différents régimes géométriques à l’équilibre thermique avant la saturation, soulignant la persistance de l’effet en présence de désordre, de sorte que l’état cohérent quantique collectif est robuste même à des températures typiques de la cellule vivante.
Comment cette étude a-t-elle été réalisée ?
L’étude a été réalisée en combinant la modélisation informatique, l’analyse théorique et les mesures expérimentales. Pour la modélisation informatique : des modèles structurels de protéines ont été créés en utilisant les coordonnées atomiques d’une ressource appelée Protein Data Bank, à partir de laquelle les coordonnées du moment dipolaire de transition des chromophores du tryptophane ont été extraites des modèles. Pour l’analyse théorique : l’optique quantique et l’approche du hamiltonien effectif (en mécanique quantique, le hamiltonien d’un système est utilisé pour calculer l’énergie totale de ce système) ont été utilisées pour décrire l’interaction entre les réseaux de tryptophane et le champ électromagnétique. Des modèles théoriques ont été développés sur la base des fonctions de corrélation dipôle-dipôle et du facteur de Boltzmann (utilisé pour calculer des éléments tels que la différence d’énergie entre deux niveaux d’énergie magnétique), à partir desquels les valeurs propres complexes de l’hamiltonien radiatif ont été résolues pour déterminer les taux de désintégration radiative et les facteurs d’augmentation du super-rayonnement. Des prédictions ont ensuite été faites pour la réponse optique collective et les rendements quantiques de fluorescence des réseaux de tryptophane.
Une fois la modélisation informatique et théorique en main, les chercheurs ont effectué les mesures expérimentales essentielles pour tester les prédictions clés basées sur les calculs. La spectroscopie à l’état stable a été réalisée sur les réseaux de tryptophane dans les architectures de protéines et les spectres d’absorption et de fluorescence ont été mesurés pour obtenir des paramètres tels que les maxima d’absorption, les maxima de fluorescence et les rendements quantiques. Les rendements quantiques mesurés ont été comparés aux prévisions théoriques afin de valider la réponse optique collective.
Les prédictions théoriques ont été comparées aux données expérimentales afin d’évaluer la précision des modèles et les effets du désordre structurel sur la réponse optique collective ont été étudiés. La robustesse des améliorations du rendement quantique au désordre statique a également été analysée. Après une analyse complète, l’équipe de recherche a pu utiliser ses données pour valider les prédictions basées sur la modélisation théorique et a démontré un effet positif du comportement quantique collectif résultant en une superradiance non triviale des chromophores de tryptophane dans les macromolécules telles que les microtubules.
L’importance du tryptophane dans le système biologique
Le tryptophane joue un rôle multiple dans la signalisation biologique [9]. Il participe à la synthèse des protéines et sert de composant clé des molécules de signalisation et des neurotransmetteurs tels que la sérotonine. Le tryptophane sert également de précurseur à la niacine et intervient dans le métabolisme énergétique et la réparation de l’ADN. Il peut agir comme chromophore, participant aux processus d’absorption et d’émission de lumière, et est utilisé comme sonde pour étudier les interactions protéine-protéine et les changements de conformation. Le tryptophane est également impliqué dans la régulation de l’expression des gènes et peut être altéré dans certaines maladies et certains troubles. Dans l’ensemble, les propriétés uniques du tryptophane en font une molécule polyvalente aux fonctions diverses dans différents processus physiologiques.
Les méga-réseaux Trp soutiennent les états quantiques dans les protéines par des effets collectifs et coopératifs. Lorsque plusieurs chromophores Trp sont organisés en réseau, leurs dipôles de transition peuvent interagir les uns avec les autres, ce qui conduit à l’émergence de modes propres superradiants et subradiants. Ces modes propres représentent des excitations collectives des chromophores Trp, où l’énergie d’excitation est partagée et distribuée dans le réseau.
Le couplage entre les dipôles de transition Trp dans les méga-réseaux permet des interactions à longue portée, même si les couplages individuels sont relativement faibles par rapport à l’énergie à température ambiante. Ce couplage à longue portée renforce la robustesse du réseau et le protège du désordre. Le fort couplage entre les chromophores Trp et le champ électromagnétique favorise la formation d’états superradiants, où le taux d’émission est considérablement amélioré par rapport à celui des chromophores Trp individuels.
Le facteur d’amélioration de la superradiation, qui compare le taux de décroissance radiative de l’état le plus brillant du réseau à celui d’un seul chromophore Trp, augmente avec la taille du réseau jusqu’à ce qu’il atteigne un point de saturation. Cela signifie que les grands méga-réseaux de Trp peuvent présenter des effets superradiants plus importants. La présence de ces états superradiants dans divers complexes biologiques, tels que les microtubules et les centrioles, est confirmée par une analyse théorique et des mesures expérimentales du rendement quantique de la fluorescence.
Ces effets optiques quantiques collectifs dans les méga-réseaux de Trp ont des implications importantes pour les systèmes biologiques. Ils peuvent améliorer l’absorption et le transfert d’énergie des photoexcitations UV, promouvoir la photoprotection en réémettant l’énergie absorbée, et potentiellement faciliter le transfert d’informations ultrarapides dans les axones neuronaux. La robustesse de ces états quantiques à l’équilibre thermique et au désordre structurel souligne leur importance potentielle dans la signalisation et le contrôle cellulaires. Il convient de noter que les molécules enrichies en fragments de tryptophane comprennent le cytochrome c des mitochondries, de sorte que cet organite est susceptible de jouer un rôle similaire et important dans le système de signalisation optique quantique de la cellule.
Principales conclusions de l’étude :
- États quantiques collectifs :
- Les chromophores Trp, lorsqu’ils sont organisés en réseaux, interagissent par l’intermédiaire de leurs dipôles de transition. Cette interaction conduit à la formation d’excitations collectives connues sous le nom de modes propres superradiants et subradiants.
- Dans les états superradiants, l’énergie d’excitation est distribuée à travers le réseau, ce qui conduit à des taux d’émission nettement plus élevés que ceux des molécules de Trp individuelles.
- Couplage à longue portée :
- Malgré les couplages individuels relativement faibles entre les dipôles de transition Trp, le réseau global présente des interactions à longue portée. Cela renforce la robustesse du réseau face au désordre structurel et aux fluctuations thermiques.
- Facteur d’amélioration du rayonnement :
- Le taux de décroissance radiative de l’état le plus brillant du réseau est beaucoup plus élevé que celui d’un chromophore Trp unique. Ce facteur d’amplification du rayonnement augmente avec la taille du réseau jusqu’à ce qu’un point de saturation soit atteint.
- Présence dans les structures biologiques :
- On trouve des états superradiants dans divers complexes biologiques, tels que les microtubules et les centrioles. Ces résultats sont étayés par une analyse théorique et des mesures expérimentales du rendement quantique de la fluorescence.
Implications pour les systèmes biologiques :
- Amélioration de l’absorption des UV et du transfert d’énergie :
- Les propriétés superradiantes des réseaux Trp améliorent l’absorption et le transfert des photoexcitations UV. Cela pourrait améliorer l’efficacité de l’utilisation de l’énergie dans les systèmes biologiques.
- Photoprotection :
- Les réseaux peuvent réémettre l’énergie absorbée, protégeant ainsi potentiellement les cellules des dommages induits par les UV en dissipant efficacement l’énergie excédentaire.
- Transfert d’informations ultrarapide :
- La robustesse et l’efficacité de ces états quantiques suggèrent un rôle potentiel dans la facilitation du transfert rapide d’informations au sein des axones neuronaux et d’autres structures cellulaires.
- Robustesse thermique et structurelle :
- Les états quantiques des méga-réseaux de Trp résistent aux fluctuations thermiques et aux désordres structurels, ce qui souligne leur importance dans le maintien des mécanismes de signalisation et de contrôle cellulaires dans des conditions variables.
L’étude démontre que le tryptophane, au-delà de ses rôles bien connus dans la synthèse des protéines, la neurotransmission et les voies métaboliques, participe également à des phénomènes d’optique quantique sophistiqués lorsqu’il est organisé en méga-réseaux. Les propriétés superradiantes de ces réseaux pourraient avoir des implications significatives pour améliorer le transfert d’énergie, la photoprotection et le traitement de l’information dans les systèmes biologiques. Ces résultats ouvrent de nouvelles voies pour comprendre les aspects quantiques des processus biologiques et leurs applications potentielles en biotechnologie et en médecine.
Les enseignements potentiels à tirer de l’étude
À partir de la modélisation théorique et de la validation expérimentale, l’étude démontre le potentiel d’effets photophysiques et photochimiques significatifs dans les méga-réseaux de tryptophane dans les architectures biologiques résultant de l’amélioration optique quantique au cours de la cohérence collective, de la résonance à longue portée et de la synchronisation. Contrairement au sentiment commun des experts selon lequel le système biologique était trop humide, bruyant et chaud pour des effets quantiques non triviaux tels que la cohérence collective, la superradiance et l’interactivité à longue portée observées, même dans l’environnement thermique ambiant de la cellule, réfutent ces perspectives naïves (naïves parce qu’elles ne tenaient pas pleinement compte des propriétés organisationnelles structurelles/géométriques uniques des protéines cellulaires et étaient basées sur des visions très simplistes de l’organisation cellulaire).
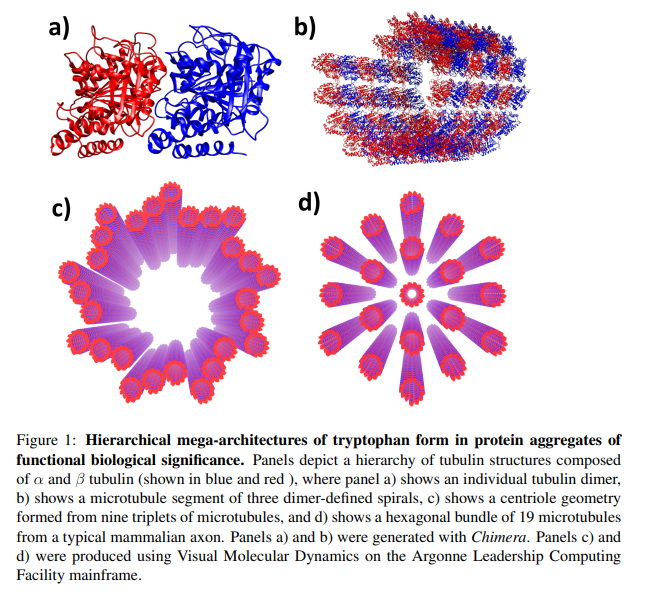
Ces résultats ont de vastes implications et sont importants car les effets optiques quantiques collectifs dans les méga-réseaux de tryptophane peuvent sous-tendre des processus clés d’orchestration et de signalisation cellulaires, comme le transfert d’informations potentiellement ultrarapide dans les axones neuronaux, qui ont des faisceaux filamentaires de microtubules hautement ordonnés (figure 2).
De telles preuves empiriques soutiennent des théories telles que le réseau unifié de mémoire spatiale de Haramein et Brown, ou la réduction objective orchestrée de Hameroff et Penrose, qui impliquent des assemblages macromoléculaires subcellulaires tels que les mitochondries et les microtubules (la matrice réticulaire mitochondriale). Par exemple, en ce qui concerne la théorie Orch-OR de Hameroff-Penrose, la physicienne Sabina Hossenfelder, commentatrice de vulgarisation scientifique, a récemment fait le commentaire suivant : « Lorsque Roger Penrose a émis l’idée que le cerveau humain utilisait des effets quantiques dans les microtubules et que c’était là l’origine de la conscience, beaucoup ont pensé que l’idée était un peu folle. Selon une nouvelle étude, il s’avère que Penrose avait raison… en tout cas en ce qui concerne les microtubules. Jetons un coup d’œil ».
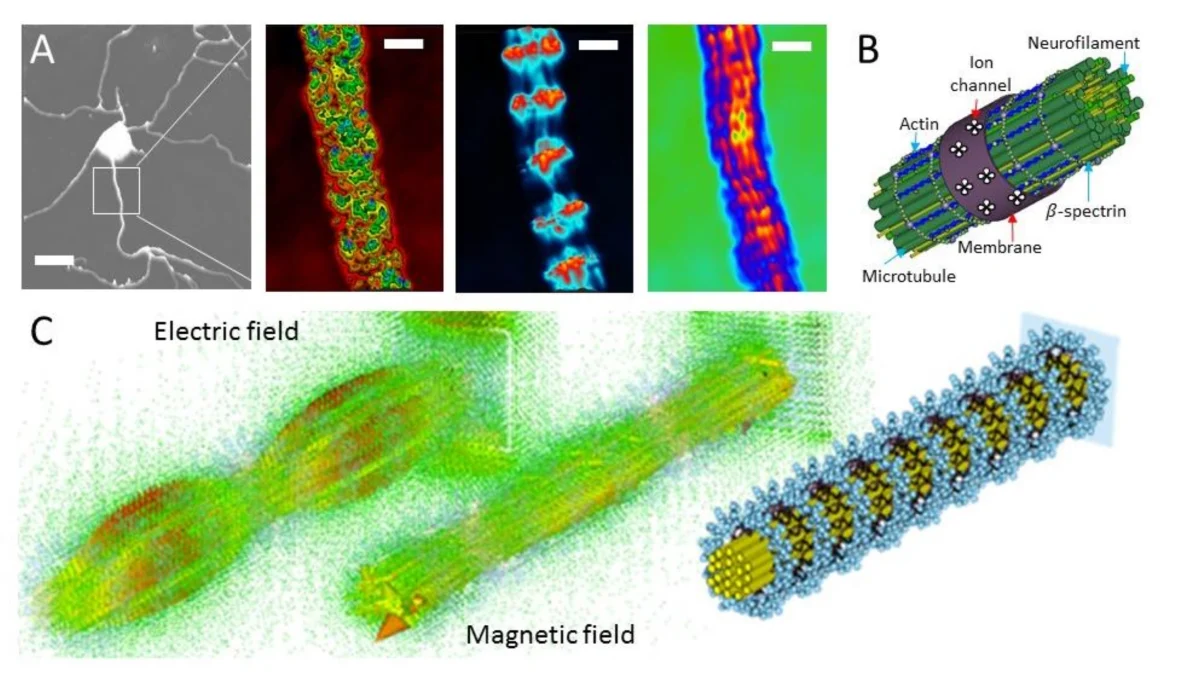
La théorie des réseaux à mémoire spatiale de Haramein et Brown évalue la cohérence collective quantique et l’intrication des dipôles des acides aminés aromatiques comme le tryptophane dans les microtubules dans le cadre de la physique unifiée de l’équivalence entre l’intrication quantique et les géométries spatio-temporelles à connexions multiples, comme décrit dans la conjecture de correspondance holographique de Maldacena-Susskind selon laquelle les ponts d’Einstein-Rosen sont égaux aux corrélations d’Einstein-Rosen-Podolsky (ERb = EPR). À partir de cette équivalence, Haramein et Brown postulent que les états collectifs quantiques de cohérence et d’intrication des dipôles électroniques dans les résidus de tubuline constituent une fonction de mémoire et que cette propriété de mémoire est le couplage de l’intrication quantique avec la géométrie multi-connexe de l’espace qui aboutit à la propriété plus générale de la mémoire spatiale (figure 3).
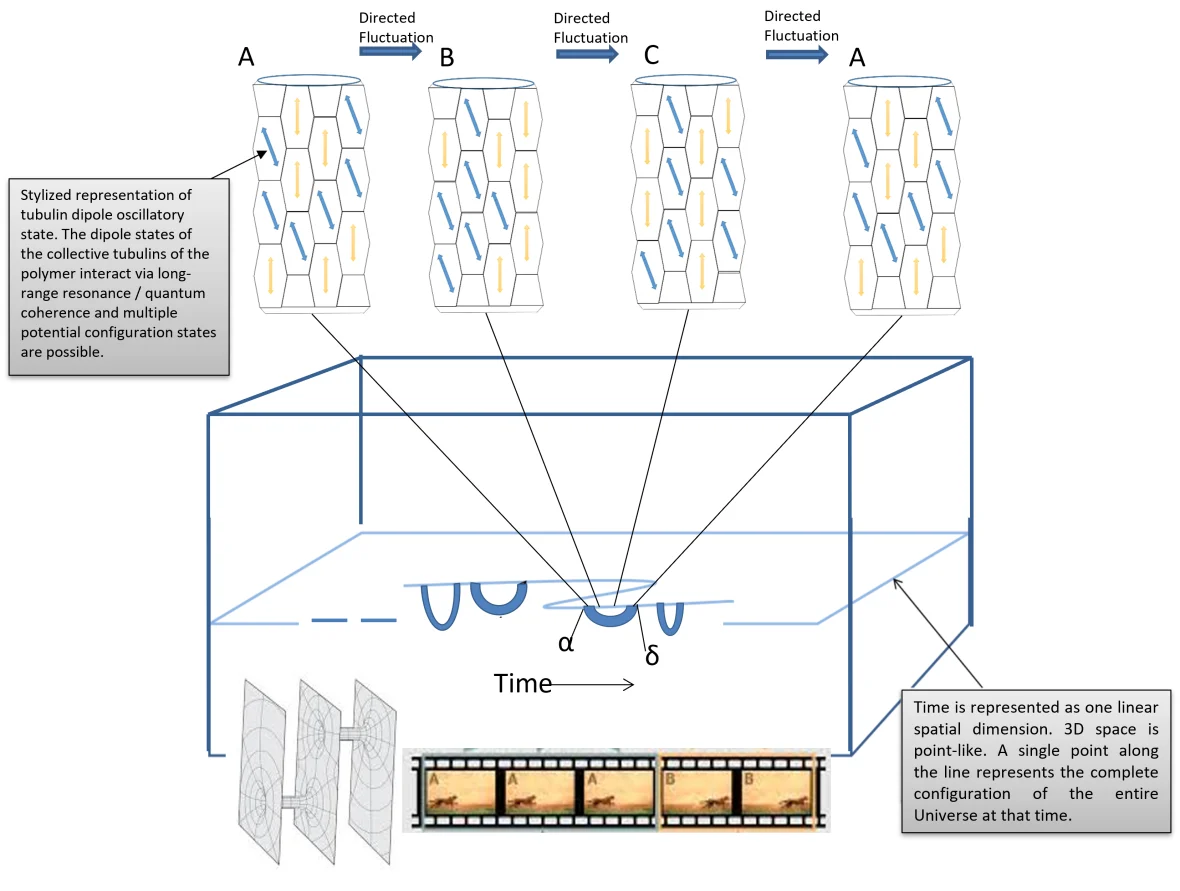
Figure 3. Image tirée de Haramein et Brown, Spacememory Theory : En raison de la forte corrélation entre l’état de configuration A des microtubules et la coordonnée spatio-temporelle α, après un certain temps au cours duquel les microtubules ont exploré plusieurs états de configuration, si la forme d’onde spatio-temporelle électromagnétique du cerveau entraîne les oscillations du dipôle électronique à proximité de la configuration A, le microtubule sera entraîné dans cette configuration, à ce moment-là, la connexion ERb de l’espace-temps avec la coordonnée α permettra l’accès à l’information de α à partir de la coordonnée de l’espace-temps δ, et donc l’information de l’état « passé » de α est disponible et accessible au moment ultérieur de δ. De cette manière, la mémoire est une adhésion quasi-instantanée de cadres spatio-temporels par le biais d’une géométrie spatiale à connexions multiples. Tous les cadres de référence spatio-temporels existent instantanément et simultanément, et c’est l’ordre du réseau d’intrication des voxels de qubits spatiaux qui ordonne une structure causale et assemble les cadres de référence spatio-temporels comme une bobine de film (en bas de l’image), qui sont connectés par le réseau ERb.
Ces deux exemples mettent en évidence l’importance des effets quantiques non triviaux dans les propriétés d’information, de cognition et éventuellement de conscience du système vivant. Cette étude la plus récente apporte une nouvelle validation à l’hypothèse théorique selon laquelle des états mécaniques quantiques non triviaux peuvent être opérationnels au sein du système vivant, une idée qui a longtemps été qualifiée de « folle » et de « farfelue » par les experts. Étant donné la remarquable intelligence sans échelle dont fait preuve le système vivant, il n’est pas surprenant que la vie ait tiré parti de phénomènes mécaniques quantiques non triviaux pour son propre bénéfice, voire qu’elle ait émergé de propriétés quantiques telles que la résonance collective à longue portée, la cohérence et la synchronisation.
Références
[1] N. S. Babcock, G. Montes-Cabrera, K. E. Oberhofer, M. Chergui, G. L. Celardo, and P. Kurian, “Ultraviolet Superradiance from Mega-Networks of Tryptophan in Biological Architectures,” J. Phys. Chem. B, vol. 128, no. 17, pp. 4035–4046, May 2024, doi: 10.1021/acs.jpcb.3c07936.
[2] N. Haramein, W. D. Brown, and A. Val Baker, “The Unified Spacememory Network: from Cosmogenesis to Consciousness,” Neuroquantology, vol. 14, no. 4, Jun. 2016, doi: 10.14704/nq.2016.14.4.961.
[3] S. Hameroff and R. Penrose, “Orchestrated reduction of quantum coherence in brain microtubules: A model for consciousness,” Mathematics and Computers in Simulation, vol. 40, no. 3, pp. 453–480, Apr. 1996, doi: 10.1016/0378-4754(96)80476-9.
[4] S. Sahu et al., “Atomic water channel controlling remarkable properties of a single brain microtubule: correlating single protein to its supramolecular assembly,” Biosens Bioelectron, vol. 47, pp. 141–148, Sep. 2013, doi: 10.1016/j.bios.2013.02.050.
[5] Frohlich, H. Long-range coherence and energy storage in biological systems. Int. J. Quantum Chem. 1968, 2, 641–649.
[6] Frohlich, H. Quantum mechanical concepts in biology. In From Theoretical Physics to Biology; Marois, M., Ed.; North-Holland: Amsterdam, the Netherlands, 1969; pp. 13–22.
[7] Frohlich, H. Long range coherence and the action of enzymes. Nature 1970, doi:10.1038/2281093a0.
[8] Vasconcellos, Áurea & Vannucchi, Fabio & Oliveira, Sergio & Luzzi, Roberto. (2012). Fröhlich Condensate: Emergence of Synergetic Dissipative Structures in Information Processing Biological and Condensed Matter Systems. Information. 3. 601-620. 10.3390/info3040601.
[9] S. Barik, “The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins,” Int J Mol Sci, vol. 21, no. 22, p. 8776, Nov. 2020, doi: 10.3390/ijms21228776.
[10] ] P. Singh et al., “Cytoskeletal Filaments Deep Inside a Neuron Are not Silent: They Regulate the Precise Timing of Nerve Spikes Using a Pair of Vortices,” Symmetry, vol. 13, no. 5, Art. no. 5, May 2021, doi: 10.3390/sym13050821.