Sinopsis
Un análisis de los efectos fotofísicos y fotoquímicos en mega-redes de triptófano en arquitecturas biológicas ha descubierto una coherencia colectiva que da lugar a una mejora óptica cuántica, conocida como superradiancia, y a una interactividad de largo alcance incluso en el entorno térmico de la célula [1]. Estos efectos ópticos cuánticos colectivos en las megarredes de triptófano pueden subyacer a procesos clave de orquestación y señalización celular, como la transferencia de información potencialmente ultrarrápida en los axones neuronales, que tienen haces filamentosos de microtúbulos centrales altamente ordenados y enriquecidos con moléculas de triptófano coordinadas que participan en el procesamiento de la información. Estas pruebas empíricas apoyan teorías como la de la Red Espacial de Memoria Unificada de Haramein y Brown [2] o la de la Reducción Objetiva Orquestada de Hameroff y Penrose [3], que implican a conjuntos macromoleculares subcelulares como las mitocondrias y los microtúbulos (la matriz reticular mitocondrial).
Lo que se Descubrió
Un equipo de investigación internacional (de EE.UU., México, Suiza e Italia) ha realizado un análisis exhaustivo que combina el modelado computacional, el análisis teórico y las mediciones experimentales para documentar y caracterizar el procesamiento colectivo de señales ópticas en mega-redes de triptófano como las que se encuentran en los microtúbulos. Por ejemplo, una superarquitectura de microtúbulos, denominada centríolo (Figura 1), de 320 nm de longitud contendrá alrededor de 105 cromóforos de triptófano (un cromóforo es una molécula que absorbe y emite luz). Con este tamaño, la resonancia de largo alcance y la sincronización de las oscilaciones dipolares convierten a las arquitecturas proteínicas como ésta en mega-redes y ayudan a explicar estudios anteriores como el de Bandopahdyay et al. [4], que han hallado una coordinación significativa entre microtúbulos individuales que se comportan como si todo el conjunto supramolecular estuviera en un estado cuántico, como un condensado de Frohlich [5-8].
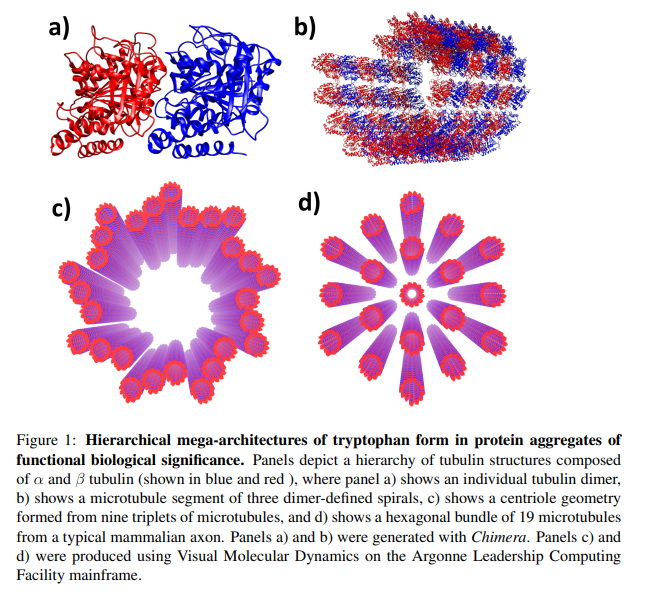
Por lo tanto, tal hallazgo apoya las teorías de que los microtúbulos ocupan papeles esenciales en el procesamiento de la información celular: posiblemente desempeñen papeles significativos en procesos cognitivos como los que implican a las neuronas. La respuesta óptica colectiva de las redes de triptófano en arquitecturas proteicas se caracteriza por los espectros de absorción y emisión de los cromóforos de la red, y el análisis de esta respuesta óptica colectiva de las redes de triptófano ha identificado mejoras superradiantes, un efecto cuántico que da lugar a una mayor intensidad de emisión del flujo electromagnético coherente, que podría ser utilizado por la célula para la señalización y el transporte de energía de alta eficiencia.
La respuesta óptica colectiva puede verse afectada por el desorden estructural, pero las redes más grandes de residuos poliméricos de triptófano coordinados examinadas en este estudio tienden a mostrar una robustez cooperativa frente al desorden. Este fenómeno implica una colección de cromóforos de triptófano idénticos que emiten espontáneamente una intensa radiación coherente. Los dipolos de transición de los cromóforos de triptófano se sincronizan coherentemente, dando lugar a estados superradiantes que mejoran la transferencia de excitación dentro de la red. Este efecto cuántico cooperativo da lugar a un pico de intensidad de emisión mayor que el de un solo cromóforo. Así pues, las propiedades fotoexcitónicas colectivas de las grandes redes de triptófano en estructuras proteicas que van desde dímeros de tubulina individuales y segmentos de microtúbulos hasta superarquitecturas de microtúbulos como el centríolo y los haces neuronales son evidentes en el rendimiento fluorescente medido, que aumenta significativamente desde tubulinas individuales hasta conjuntos de tubulinas, una característica clave de la superradiancia y, por tanto, de las propiedades cuánticas coherentes colectivas.
La respuesta óptica colectiva está influida por el acoplamiento entre los cromóforos del triptófano inducido por el campo electromagnético. La respuesta óptica colectiva de las redes de triptófano en arquitecturas proteicas se caracteriza por la mejora del rendimiento cuántico (QY). El QY es una cifra de mérito para la cooperatividad y es una medida robusta de la mejora de la respuesta óptica. El QY no se ve afectado significativamente por el desorden estático, incluso para fuerzas de desorden de la misma magnitud que la energía a temperatura ambiente. Esta robustez se atribuye a la distribución de la fuerza del dipolo superradiante entre otros estados excitónicos, mientras que los estados cercanos al estado superradiante en energía siguen exhibiendo la mayor parte de la fuerza del dipolo.
Por tanto, la respuesta óptica colectiva de las redes de triptófano en arquitecturas proteicas se caracteriza por una mejora persistente del rendimiento cuántico, incluso en condiciones ambientales. La respuesta óptica colectiva de las redes de triptófano en arquitecturas proteicas es la formación de estados fuertemente superradiantes debido a interacciones colectivas entre disposiciones organizadas de hasta más de 105 dipolos de transición excitados por UV de triptófano en arquitecturas de microtúbulos. Esto conduce a un aumento del rendimiento cuántico de fluorescencia, que ha sido confirmado por experimentos. El comportamiento superradiante en el rendimiento cuántico de fluorescencia se observa en estructuras de tubulina organizadas jerárquicamente, como microtúbulos, centriolos y haces neuronales. El rendimiento cuántico aumenta en diferentes regímenes geométricos en equilibrio térmico antes de la saturación, lo que pone de relieve la persistencia del efecto en presencia de desorden, de modo que el estado coherente cuántico colectivo es robusto incluso a temperaturas típicas de la célula viva.
¿Cómo se Realizó este Estudio?
El estudio se llevó a cabo mediante una combinación de modelización computacional, análisis teórico y mediciones experimentales. Para el modelado computacional: se crearon modelos estructurales de proteínas utilizando coordenadas atómicas de un recurso llamado Banco de Datos de Proteínas, del que se extrajeron las coordenadas del momento dipolar de transición de los cromóforos del triptófano. Para el análisis teórico: se utilizaron enfoques de óptica cuántica y hamiltoniano efectivo (en mecánica cuántica, el hamiltoniano de un sistema se utiliza para calcular la energía total de ese sistema) para describir la interacción entre las redes de triptófano y el campo electromagnético. Se desarrollaron modelos teóricos basados en las funciones de correlación dipolo-dipolo y el factor de Boltzmann (utilizado para calcular cosas como la diferencia de energía entre dos niveles de energía magnética), a partir de los cuales se resolvieron los valores propios complejos del hamiltoniano radiativo para determinar las tasas de desintegración radiativa y los factores de aumento superradiante. A continuación, se hicieron predicciones para la respuesta óptica colectiva y los rendimientos cuánticos de fluorescencia de las redes de triptófano.
Con los modelos informáticos y teóricos a mano, los investigadores realizaron las importantísimas mediciones experimentales para comprobar las predicciones clave basadas en los cálculos. Se realizó espectroscopia de estado estacionario en redes de triptófano en arquitecturas proteicas y se midieron los espectros de absorción y fluorescencia para obtener parámetros como los máximos de absorción, los máximos de fluorescencia y los rendimientos cuánticos. Los rendimientos cuánticos medidos se compararon con las predicciones teóricas para validar la respuesta óptica colectiva.
Las predicciones teóricas se compararon con datos experimentales para evaluar la precisión de los modelos y se investigaron los efectos del desorden estructural en la respuesta óptica colectiva. También se analizó la solidez de las mejoras del rendimiento cuántico frente al desorden estático. Tras un análisis exhaustivo, el equipo de investigación pudo utilizar sus datos para validar las predicciones basadas en el modelado teórico y demostró un efecto positivo del comportamiento cuántico colectivo que resulta en una superradiancia no trivial de los cromóforos de triptófano en macromoléculas como los microtúbulos.
La Importancia del Triptófano en el Sistema Biológico
El triptófano desempeña un papel polifacético en la señalización biológica [9]. Participa en la síntesis de proteínas y es un componente clave de moléculas de señalización y neurotransmisores como la serotonina. El triptófano también actúa como precursor de la niacina y participa en el metabolismo energético y la reparación del ADN. Puede actuar como cromóforo, participando en procesos de absorción y emisión de luz, y se utiliza como sonda para estudiar las interacciones proteína-proteína y los cambios conformacionales. El triptófano también interviene en la regulación de la expresión génica y puede verse alterado en determinadas enfermedades y trastornos. En general, las propiedades únicas del triptófano lo convierten en una molécula versátil con diversas funciones en varios procesos fisiológicos.
Las mega-redes de Trp apoyan los estados cuánticos en las proteínas a través de efectos colectivos y cooperativos. Cuando múltiples cromóforos Trp se organizan en una red, sus dipolos de transición pueden interactuar entre sí, dando lugar a la aparición de modos propios superradiantes y subradiantes. Estos modos propios representan excitaciones colectivas de los cromóforos Trp, en las que la energía de excitación se comparte y distribuye entre la red.
El acoplamiento entre los dipolos de transición Trp en las mega-redes permite interacciones de largo alcance, aunque los acoplamientos individuales son relativamente débiles en comparación con la energía a temperatura ambiente. Este acoplamiento de largo alcance aumenta la robustez de la red y la protege del desorden. El fuerte acoplamiento entre los cromóforos Trp y el campo electromagnético favorece la formación de estados superradiantes, en los que la tasa de emisión aumenta significativamente en comparación con la de los cromóforos Trp individuales.
El factor de mejora superradiante, que compara la tasa de desintegración radiativa del estado más brillante de la red con la de un único cromóforo Trp, aumenta con el tamaño de la red hasta alcanzar un punto de saturación. Esto significa que las mega-redes de Trp más grandes pueden mostrar efectos superradiantes más fuertes. La presencia de estos estados superradiantes en diversos complejos biológicos, como microtúbulos y centríolos, se confirma mediante análisis teóricos y mediciones experimentales del rendimiento cuántico de fluorescencia.
Estos efectos ópticos cuánticos colectivos en las mega-redes de Trp tienen importantes implicaciones para los sistemas biológicos. Pueden mejorar la absorción y la transferencia de energía de las fotoexcitaciones UV, promover la fotoprotección mediante la reemisión de la energía absorbida y facilitar potencialmente la transferencia ultrarrápida de información en los axones neuronales. La robustez de estos estados cuánticos frente al equilibrio térmico y el desorden estructural subraya aún más su importancia potencial en la señalización y el control celulares. Cabe señalar que entre las moléculas enriquecidas en moléculas de triptófano se encuentra el citocromo c de la mitocondria, por lo que es probable que este orgánulo desempeñe un papel similar e importante en el sistema de señalización óptica cuántica de la célula.
Conclusiones clave del estudio:
- Estados cuánticos colectivos:
- Los cromóforos Trp, cuando se organizan en redes, interactúan a través de sus dipolos de transición. Esta interacción conduce a la formación de excitaciones colectivas conocidas como modos propios superradiante y subradiante.
- En los estados superradiantes, la energía de excitación se distribuye por toda la red, lo que da lugar a tasas de emisión significativamente mayores en comparación con las moléculas individuales de Trp.
2. Acoplamiento de largo alcance:
- A pesar de los acoplamientos individuales relativamente débiles entre los dipolos de transición Trp, la red global presenta interacciones de largo alcance. Esto aumenta la robustez de la red frente al desorden estructural y las fluctuaciones térmicas.
3. Factor de Superradiancia amplificado:
- La tasa de desintegración radiativa del estado más brillante de la red es mucho mayor que la de un único cromóforo Trp. Esta mejora superradiante aumenta con el tamaño de la red hasta que se alcanza un punto de saturación.
4. Presencia en estructuras biológicas:
- Los estados superradiantes se encuentran en diversos complejos biológicos, como los microtúbulos y los centríolos. Estos hallazgos se apoyan en análisis teóricos y mediciones experimentales del rendimiento cuántico de fluorescencia.
Implicaciones para los sistemas biológicos:
- Absorción y transferencia de energía UV mejoradas:
- Las propiedades superradiantes de las redes de Trp mejoran la absorción y transferencia de fotoexcitaciones UV. Esto podría mejorar la eficiencia de la utilización de la energía en los sistemas biológicos.
2. Fotoprotección:
- Las redes pueden reemitir la energía absorbida, protegiendo potencialmente a las células de los daños inducidos por los rayos UV al disipar eficazmente el exceso de energía.
3. Transferencia ultrarrápida de información:
- La robustez y eficacia de estos estados cuánticos sugieren un papel potencial en la facilitación de la transferencia rápida de información dentro de los axones neuronales y otras estructuras celulares.
4. Robustez Térmica y estructural
- Los estados cuánticos de las megarredes de Trp son resistentes a las fluctuaciones térmicas y al desorden estructural, lo que pone de relieve su importancia para mantener los mecanismos de señalización y control celular en condiciones variables.
El estudio demuestra que el triptófano, más allá de sus conocidas funciones en la síntesis de proteínas, la neurotransmisión y las vías metabólicas, también participa en sofisticados fenómenos ópticos cuánticos cuando se organiza en megarredes. Las propiedades superradiantes de estas redes podrían tener importantes implicaciones para mejorar la transferencia de energía, la fotoprotección y el procesamiento de la información en los sistemas biológicos. Los hallazgos abren nuevas vías para comprender los aspectos cuánticos de los procesos biológicos y sus posibles aplicaciones en biotecnología y medicina.
Posibles Conclusiones del Estudio
A partir de la modelización teórica y la validación experimental, el estudio demuestra el potencial de importantes efectos fotofísicos y fotoquímicos en megarredes de triptófano en arquitecturas biológicas resultantes de la mejora óptica cuántica durante la coherencia colectiva, la resonancia de largo alcance y la sincronización. Contrariamente a lo que solía ser el sentimiento común entre los expertos de que el sistema biológico era demasiado húmedo, ruidoso y cálido para efectos cuánticos no triviales como la coherencia colectiva, la superradiancia observada y la interactividad de largo alcance incluso en el entorno térmico ambiental de la célula refutan tales perspectivas ingenuas (ingenuas porque no tenían plenamente en cuenta las propiedades organizativas estructurales / geométricas únicas de las proteínas celulares y se basaban en visiones muy simplistas de la organización celular).
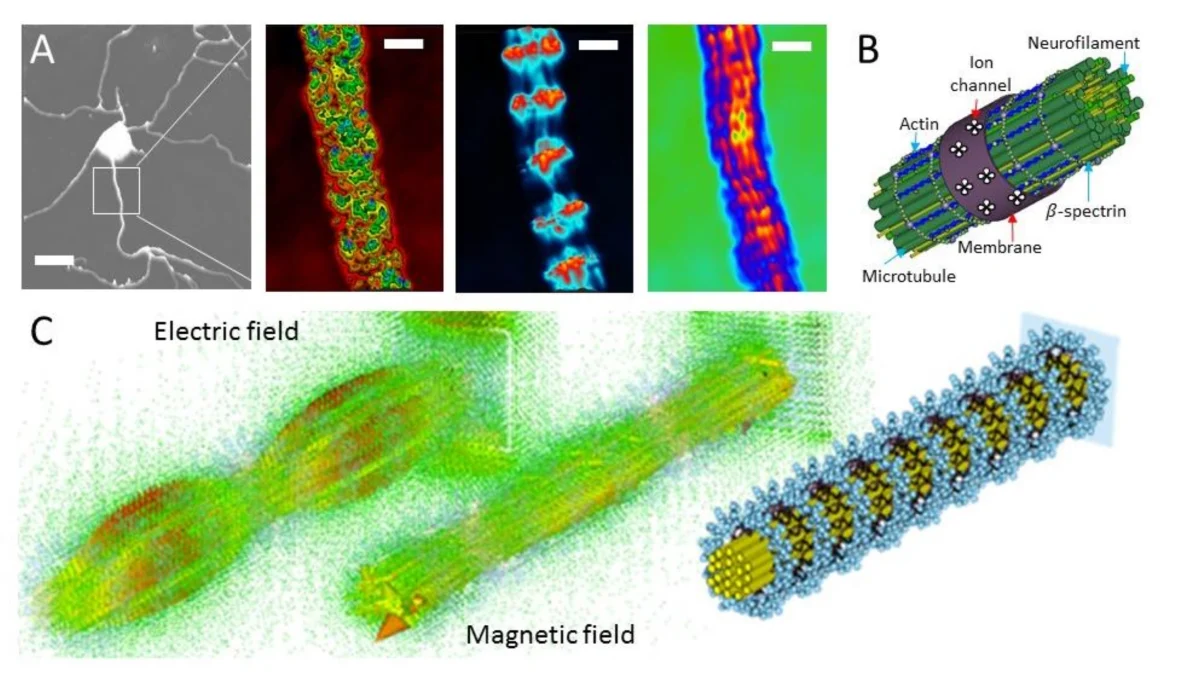
Los hallazgos tienen implicaciones de gran alcance y son importantes porque los efectos ópticos cuánticos colectivos en las mega-redes de triptófano pueden subyacer a procesos clave de orquestación y señalización celular, como la transferencia de información potencialmente ultrarrápida en los axones neuronales, que tienen haces filamentosos de microtúbulos centrales altamente ordenados (Figura 2).
Tales pruebas empíricas apoyan teorías como La Red Espacial de Memoria Unificada de Haramein y Brown, o la Reducción Objetiva Orquestada de Hameroff y Penrose, que implican a conjuntos macromoleculares subcelulares como las mitocondrias y los microtúbulos (la matriz reticular mitocondrial). Como ejemplo, en relación con la teoría Orch-OR de Hameroff-Penrose, la física Sabina Hossenfelder, comentarista de divulgación científica, comentó recientemente: «Cuando Roger Penrose propuso originalmente la idea de que el cerebro humano utiliza efectos cuánticos en los microtúbulos y que ése era el origen de la conciencia, muchos pensaron que la idea era un poco descabellada. Según un nuevo estudio, resulta que Penrose tenía razón… en lo que respecta a los microtúbulos. Echemos un vistazo».
La Teoría de la Red Espacial de Memoria de Haramein y Brown evalúa la coherencia colectiva cuántica y el entrelazamiento de los dipolos de los aminoácidos aromáticos como el triptófano en los microtúbulos dentro del marco de la Física Unificada de equivalencia entre el entrelazamiento cuántico y las geometrías espaciotemporales de conexión múltiple, como se describe en la conjetura de correspondencia holográfica de Maldacena-Susskind de que los puentes de Einstein-Rosen son iguales a las correlaciones de Einstein-Rosen-Podolsky (ERb = EPR). A partir de esta equivalencia, Haramein y Brown postulan que los estados colectivos cuánticos coherentes y de entrelazamiento de los dipolos electrónicos en los residuos de tubulina una función en la memoria y esta propiedad de memoria es el acoplamiento del entrelazamiento cuántico con la geometría de múltiples conexiones del espacio que da lugar a la propiedad más general de la Espacimemoria (Figura 3).
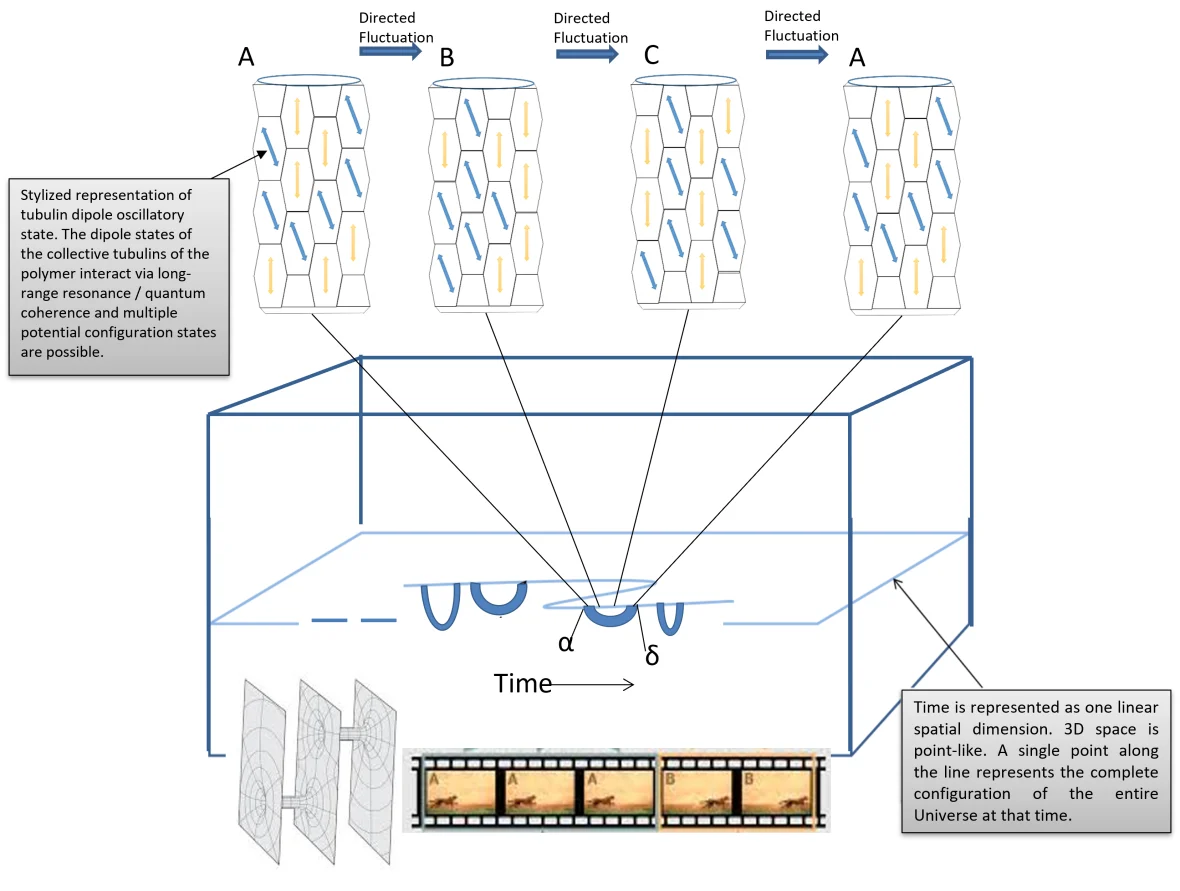
Figura 3 Imagen de Haramein y Brown, Spacememory Theory: debido a la fuerte correlación del estado de configuración A del microtúbulo con la coordenada α del espaciotiempo, después de un tiempo transcurrido en el que el microtúbulo ha explorado múltiples estados de configuración, si la forma de onda electromagnética espaciotemporal del cerebro impulsa las oscilaciones del dipolo electrónico cerca de la configuración A, será impulsado a esta configuración, en cuyo momento la conexión espaciotemporal ERb con la coordenada α permitirá entonces la adhesión de información de α desde la coordenada espaciotemporal δ, y así la información del estado «pasado» de α está disponible accesible en el momento posterior de δ. De este modo, la memoria es una adhesión casi instantánea de los marcos espaciotemporales a través de una geometría espacial de conexión múltiple. Todos los marcos de referencia espaciotemporales existen instantáneamente de forma simultánea, y es la ordenación de la red de entrelazamiento de los vóxeles qubit espaciales la que ordena una estructura causal y recompone los marcos de referencia espaciotemporales como un carrete de película (parte inferior de la imagen), que están conectados por la red ERb.
De ambos ejemplos se desprende la importancia de los efectos cuánticos no triviales en las propiedades informativas, cognitivas y, posiblemente, de conciencia del sistema vivo. Este último estudio corrobora la hipótesis teórica de que los estados mecánicos cuánticos no triviales pueden ser operativos en el sistema vivo, una idea que durante mucho tiempo fue tachada de «loca» y «chiflada» por los expertos. Dada la notable inteligencia sin escalas que exhibe el sistema vivo, no debería sorprender que la vida haya aprovechado fenómenos mecánicos cuánticos no triviales en su propio beneficio, o incluso que posiblemente haya surgido de propiedades cuánticas como la resonancia colectiva de largo alcance, la coherencia y la sincronización.
Referencias
[1] N. S. Babcock, G. Montes-Cabrera, K. E. Oberhofer, M. Chergui, G. L. Celardo, and P. Kurian, “Ultraviolet Superradiance from Mega-Networks of Tryptophan in Biological Architectures,” J. Phys. Chem. B, vol. 128, no. 17, pp. 4035–4046, May 2024, doi: 10.1021/acs.jpcb.3c07936.
[2] N. Haramein, W. D. Brown, and A. Val Baker, “The Unified Spacememory Network: from Cosmogenesis to Consciousness,” Neuroquantology, vol. 14, no. 4, Jun. 2016, doi: 10.14704/nq.2016.14.4.961.
[3] S. Hameroff and R. Penrose, “Orchestrated reduction of quantum coherence in brain microtubules: A model for consciousness,” Mathematics and Computers in Simulation, vol. 40, no. 3, pp. 453–480, Apr. 1996, doi: 10.1016/0378-4754(96)80476-9.
[4] S. Sahu et al., “Atomic water channel controlling remarkable properties of a single brain microtubule: correlating single protein to its supramolecular assembly,” Biosens Bioelectron, vol. 47, pp. 141–148, Sep. 2013, doi: 10.1016/j.bios.2013.02.050.
[5] Frohlich, H. Long-range coherence and energy storage in biological systems. Int. J. Quantum Chem. 1968, 2, 641–649.
[6] Frohlich, H. Quantum mechanical concepts in biology. In From Theoretical Physics to Biology; Marois, M., Ed.; North-Holland: Amsterdam, the Netherlands, 1969; pp. 13–22.
[7] Frohlich, H. Long range coherence and the action of enzymes. Nature 1970, doi:10.1038/2281093a0.
[8] Vasconcellos, Áurea & Vannucchi, Fabio & Oliveira, Sergio & Luzzi, Roberto. (2012). Fröhlich Condensate: Emergence of Synergetic Dissipative Structures in Information Processing Biological and Condensed Matter Systems. Information. 3. 601-620. 10.3390/info3040601.
[9] S. Barik, “The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins,” Int J Mol Sci, vol. 21, no. 22, p. 8776, Nov. 2020, doi: 10.3390/ijms21228776.
[10] ] P. Singh et al., “Cytoskeletal Filaments Deep Inside a Neuron Are not Silent: They Regulate the Precise Timing of Nerve Spikes Using a Pair of Vortices,” Symmetry, vol. 13, no. 5, Art. no. 5, May 2021, doi: 10.3390/sym13050821.


