Introduction
When we think about biology, we usually picture chemistry: molecules bumping into each other, enzymes reacting, and signals spreading by diffusion. That picture is real—but it may be incomplete. In my recent paper in Harmonic Science Perspectives (Vol 1, Issue 1), I propose a complementary layer of cellular organization: a fast, coordination-capable “resonance network” that uses three interchangeable carriers of energy and information.
Those three carriers are light (photons), vibration/sound-like mechanical waves (phonons), and mobile electronic excitations in biomolecules (excitons). The central idea is simple to state even if the details are deep: living systems may continuously convert energy back and forth between these three modes to synchronize activity across space and time inside the cell—and potentially across tissues.
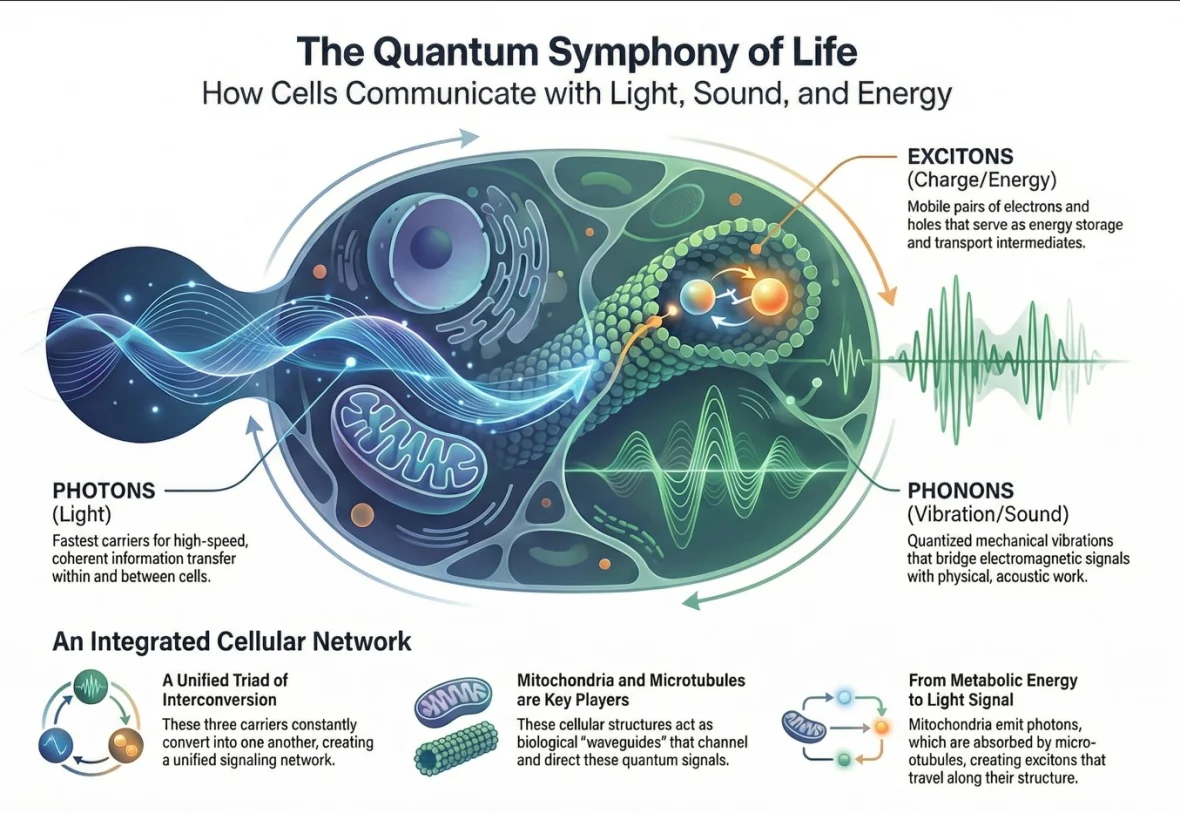
A simple picture: three messengers that can translate into one another
Imagine a cell as a city that needs rapid coordination. Chemical signals are like delivery trucks: powerful, specific, but relatively slow and locally constrained. The photon-phonon-exciton framework proposes that cells also have something more like a high-speed communication layer—where messages can move as light pulses, mechanical vibrations, or electronic excitations, and then be translated from one form into another depending on what the cell needs.
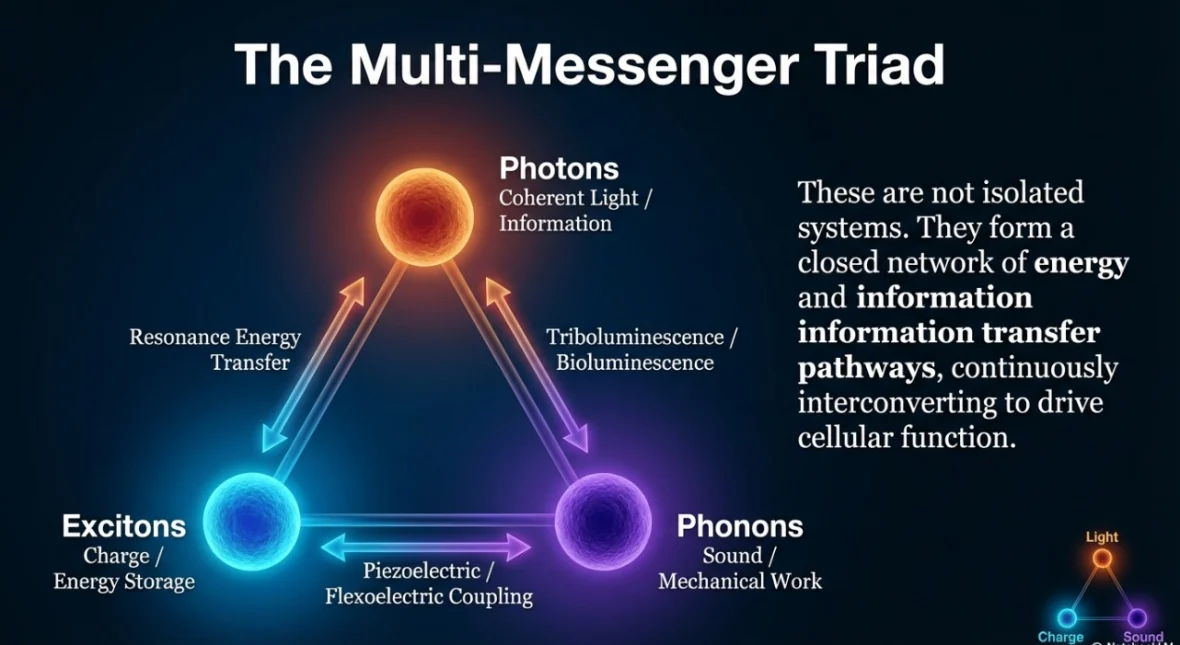
In the paper, I describe this as a “triad”:
Photons (light): Photons are extremely fast and can, in principle, carry timing information. Cells also emit ultra-weak light as part of normal metabolism, especially through mitochondrial processes linked to reactive oxygen species (ROS). This is not bioluminescence like a firefly—it’s far subtler—but it’s measurable and increasingly studied.
Phonons (vibrations, including sound-like waves): Phonons are the quantum description of mechanical vibrations. In biology, that can look like pressure waves, vibrational energy moving through proteins, or coherent mechanical pulses associated with cellular activity. Mechanical signaling is not just “support structure” in the cell; it can be a real information channel.
Excitons (mobile electronic excitations): An exciton is a bound electron-hole excitation that can move through a material. In biological contexts, exciton-like behavior can occur in aromatic or pigment-rich networks—places where electrons can become delocalized and energy can migrate efficiently. This is already a well-established concept in photosynthesis, and there is growing interest in whether related phenomena exist in other biomolecular structures as well.
Where this shows up in the body: mitochondria and microtubules as a coupled network
Two structures show up repeatedly in the evidence and the theoretical synthesis: mitochondria and microtubules.
Mitochondria as metabolic light sources. Mitochondria don’t just make ATP. Their electron transport processes can produce electronically excited intermediates, and when those relax, they can emit photons—often discussed under the umbrella of ultra-weak photon emission (UPE). These emissions can carry information about metabolic state, oxidative stress, and cellular redox balance.

Microtubules as potential waveguides and transducers. Microtubules are famous for their role in cell structure and intracellular transport. But they also have intriguing physical properties: they are highly ordered polymers, rich in aromatic amino acids (like tryptophan and tyrosine), and they can plausibly couple mechanical vibrations to electrical effects (as described in the broader literature). The hypothesis is not that microtubules “replace” chemistry, but that they may provide a physical infrastructure that supports rapid coordination: absorbing photons, launching excitons, and coupling into mechanical modes.

A concrete example discussed in the paper is: mitochondria emit ultra-weak photons; microtubule chromophores absorb them; excitons migrate along the microtubule lattice; and recombination can re-emit photons, potentially supporting signaling and synchronization.
Why interconversion matters: translation is the key feature
The strongest claim in the framework is not merely that these three phenomena exist (they do, in various forms), but that their ability to translate into one another could be what makes them biologically powerful.
Photon to exciton: light absorbed into electronic excitation. Exciton to phonon: electronic excitation couples into vibration (or vice versa). Phonon to photon: vibrational systems can influence emission and absorption dynamics under certain conditions.
This matters because each carrier has different strengths. Light is fast. Vibrations couple naturally to structure and force. Excitons can store and route energy through specific molecular networks. A cell that can move information across these channels gains a kind of multi-modal coordination system rather than relying on one mode alone.
Resonant synchronization: a possible mechanism for cellular timing
Cells don’t just need signals—they need timing. In the paper I propose that the triad can support phase-locking: stable synchronization among optical, mechanical, and electronic oscillations. In plain language, phase-locking means different oscillators “keep time” together, resisting noise and drift.
To understand what phase-locking means, consider fireflies in a forest. Each firefly has its own natural rhythm, blinking at its own pace. But under the right conditions, something remarkable happens: thousands of fireflies begin to flash in unison. When they lock together, the effect becomes unmistakable and powerful—their individual signals amplify each other through coordination. What was random noise becomes a coherent beacon. This is phase-locking: independent oscillators finding and maintaining a common rhythm.
This phenomenon isn’t just poetic – it’s already critical to how your body works. Consider neural gamma oscillations in the brain. During attention and memory tasks, different brain regions phase-lock their electrical activity, synchronizing their rhythms in the 30-100 Hz range. When this synchronization is strong, cognition is sharp. When it breaks down – whether through disease, fatigue, or disruption—cognitive performance suffers. The brain regions are still active, still firing, but they’ve lost their coordinated timing. The orchestra is playing, but no longer together.
If such phase-locking exists more broadly in living systems—not just in neural electrical activity, but across optical, mechanical, and electronic oscillations within cells—it could help explain how cells maintain coordinated rhythms across multiple scales. From fast molecular events to slower cellular and tissue-level dynamics, timing could be maintained without requiring everything to be driven purely by slow diffusion and chance encounters.
Therapeutic implications: why light and sound therapies might work better together
One motivation for exploring this framework is that modern medicine is already using parts of it—often without a unifying theory.
Photobiomodulation (PBM), for example, uses red and near-infrared light to influence mitochondrial function and cellular metabolism. Separately, low-intensity ultrasound and other mechanotherapies can influence tissue repair, signaling, and structure. The paper sketches the idea of “quantum physiotherapy”: deliberately co-tuning light-based stimulation with low-intensity ultrasound so the system is encouraged to move coherently across the photon-phonon-exciton triangle, rather than stimulating only one corner of it.
A related concept for neuromodulation is combining magnetic stimulation with acoustic entrainment in frequency ranges associated with brain rhythms. The key point is not a clinical claim, but a research direction: if biology uses coupled oscillations, then multi-modal stimulation may be a more “native language” for the nervous system than single-modality inputs.
A note on what’s established vs what’s proposed
It’s important to be clear about scientific status. Many of the components discussed are empirically supported fields of research (ultra-weak photon emission, mechanobiology, excitonic coherence in photosynthesis, and advanced imaging of cellular mechanical modes, among others). The triad as a unified organizing principle – and especially the stronger ideas about deep coherence and system-wide synchronization—are integrative hypotheses. They aim to connect real observations into a testable framework and motivate new experiments.
The paper closes with a practical experimental attitude: if the triad is real as a coupled system, then we should be able to measure signatures of coordinated timing across photon emission, phonon propagation, and exciton dynamics – ideally in the same cellular structures with integrated instrumentation.
Conclusion: a new lens on living organization
The central message is that biology may be more than chemistry plus heat and noise. Life might also be, in part, an orchestration of coherent energy flows—light, vibration, and electronic excitation—interconverting in a way that supports coordination and resilience.

If that sounds unusual, it’s worth remembering: the history of science is full of moments where a new measurement capability reveals a hidden layer of nature. Today, advances in ultrafast spectroscopy, single-photon imaging, and high-resolution mechanical mapping are making it possible to ask questions about living systems that were not experimentally accessible before. My hope is that the photon-phonon-exciton framework gives a clear conceptual map for what to look for next – and why it could matter for health, cognition, and the future of biophysics.
Check out the published article in the premiere issue of Harmonic Science Perspectives (link below):



